Inmunología Humana
-
INTRODUCCIÓN AL SISTEMA INMUNE HUMANOIntroducción. Conceptos básicos10 Temas
-
Introducción Inmunología
-
Funciones sistema inmune y barreras de defensa
-
Inmunidad innata vs adaptativa
-
Respuesta humoral vs celular
-
Respuesta adaptativa primaria vs secundaria
-
Características de la respuesta inmunitaria adaptativa
-
Concepto de antígeno
-
La respuesta inmunitaria específica es clonal
-
Revisión de los componentes del sistema inmunitario
-
Patología general del sistema inmunitario
-
Introducción Inmunología
-
Células del sistema inmune y diferenciación celular6 Temas
-
Hematopoyesis
-
Las células sanguíneas (I): Granulocitos y Mastocitos
-
Las células sanguíneas (II): Plaquetas, Eritrocitos, Monocitos y Macrófagos
-
Las células sanguíneas (III): Linfocitos (T y B), Células plasmáticas y LGL (Linfocitos NK)
-
Las células sanguíneas (IV): Células dendríticas y proporciones relativas de los tipos celulares
-
Funciones y gestión de receptores para antígenos
-
Hematopoyesis
-
Tejidos del sistema inmune: órganos linfoides 1º y 2º3 Temas
-
Células y mecanismos de la inmunidad innata (I): macrófagos, receptores y mecanismos efectores5 Temas
-
Células y mecanismos de la inmunidad innata (II): linfocitos NK, receptores y mecanismos efectores4 Temas
-
MOLÉCULAS IMPLICADAS EN EL RECONOCIMIENTO DE ANTÍGENOEl receptor de antígeno del linfocito B6 Temas
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
Inmunoglobulinas (II): Clases, formas, variabilidad y tejidos
-
Inmunoglobulinas (III): Enlaces antígeno-anticuerpo y funciones
-
Inmunoglobulinas (IV): Receptores para inmunoglobulinas
-
Inmunoglobulinas (V): Procesamiento de RNA, receptor del linfocito B, complejo co-receptor y activación célula B
-
Inmunoglobulinas (VI): Desarrollo / Diferenciación células B
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
El receptor de antígeno del linfocito T4 Temas
-
Mecanismos de generación de la diversidad de linfocitos T y B9 Temas
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
Reordenamiento de los genes
-
Mecanismos de amplificación de la diversidad
-
Expresión de las inmunoglobulinas en el BCR
-
Maduración de los linfocitos B
-
La cooperación T-B
-
Anomalías/Problemas en el proceso (CLÍNICA)
-
Maduración de los linfocitos B
-
Variabilidad en el repertorio del TcR en linfocitos T
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
El complejo principal de histocompatibilidad (I): estructura proteica, genética y nomenclatura3 Temas
-
El complejo principal de histocompatibilidad (II): Procesamiento y presentación de antígeno, polimorfismo y aplicaciones clínicas5 Temas
-
MOLÉCULAS ACCESORIAS DE LA RESPUESTA INMUNEEl sistema del complemento y sus receptores (I): vía clásica y vía alternativa4 Temas
-
El sistema del complemento y sus receptores (II): vía de las lectinas, vía lítica y regulación3 Temas
-
Moléculas implicadas en la comunicación intercelular (I): citocinas y sus receptores5 Temas
-
Moléculas implicadas en la comunicación intercelular (II): moléculas de adhesión y sus ligandos3 Temas
-
EL SISTEMA INMUNE EN ACCIÓN BLOQUEGeneración de linfocitos T efectores4 Temas
-
Generación de linfocitos B efectores7 Temas
-
Sistema Inmune asociado a mucosas (MALT)9 Temas
-
Introducción MALT
-
Estructura del tejido linfoide asociado a la mucosa intestinal
-
Recirculación y migración selectiva de linfocitos al intestino
-
Barrera epitelial intestinal: Función Inmunológica
-
Receptores para el reconocimiento de patrones moleculares
-
Inmunoglobulinas Poliméricas: IgM + IgA
-
Vías de entrada del antígeno en el intestino
-
Diferenciación de los linfocitos T efectores MALT
-
Tolerancia Oral ante antígenos solubles
-
Introducción MALT
-
La respuesta inmune (I): inmunidad innata e inflamación aguda8 Temas
-
Barreras no específicas frente a infección por microorganismos patógenos
-
Principales leucocitos que paticipan en las inmunidad innata
-
La rección inflamatoria
-
Inflamación aguda
-
Sucesos del proceso inflamatorio
-
Citocinas producidas por macrófagos: efectos locales y sistémicos
-
Efectos de las citocinas en la inmunidad innata
-
Proteínas de fase aguda
-
Barreras no específicas frente a infección por microorganismos patógenos
-
La respuesta inmune (II): mecanismos de la inmunidad específica8 Temas
-
La respuesta inmune (III): respuesta frente a virus, bacterias y hongos, protozoos y helmintos9 Temas
-
Principales enfermedades infecciosas mortales
-
Patógenos infecciosos
-
Respuestas frente a bacterias extracelulares
-
Respuesta frente a bacterias intracelulares
-
Proceso de fagocitosis
-
Respuestas frente a protozoos y helmintos
-
Ciclo vital del plasmodium
-
Respuestas inmunitarias frente a virus
-
Mecanismos de evasión inmunitaria. Virales y bacterianos.
-
Principales enfermedades infecciosas mortales
-
REGULACIÓN e INTRODUCCIÓN A LA INMUNOPATOLOGÍARegulación de la respuesta inmune (I): regulación por moléculas8 Temas
-
Regulación de la respuesta inmune: Introducción
-
Regulación por moléculas: el complemento (I)
-
Proteínas reguladoras del complemento y proceso de regulación (II)
-
Proceso de regulación por complemento (III)
-
Regulación de los anticuerpos preformados
-
Regulación por anticuerpos: Redes idiotípicas
-
Regulación por moléculas de linfocitos T
-
Regulación a nivel bioquímico: ITAM frente a ITIM
-
Regulación de la respuesta inmune: Introducción
-
Regulación de la respuesta inmune (II): regulación por células y sistemas4 Temas
-
El sistema inmune a lo largo del ciclo vital: Inmunosenescencia6 Temas
-
Introducción a la inmunopatología13 Temas
-
Conceptos básicos
-
Inmunodeficiencias
-
Las Inmunodeficiencias (II): Déficits de Complemento
-
Las Inmunodeficiencias (III): Déficits de citocinas en receptores
-
Las Inmunodeficiencias (IV): Déficits de TLR's
-
Las Inmunodeficiencias (V): Déficits de función fagocitaria
-
Las Inmunodeficiencias (VI): Déficit de linfocitos B
-
Las Inmunodeficiencias (VII): Déficits de linfocitos T
-
Inmunodeficiencias primarias (IDP's): Clínica y Diagnóstico
-
Hipersensibilidades
-
Autoinmunidad
-
Trasplantes. Rechazo
-
Inmunidad frente a tumores
-
Conceptos básicos
-
Introducción a la Inmunoterapia8 Temas
-
Vacunas (I): Bases inmunológicas
-
Vacunas (II): Tipos de vacunas
-
Inmunoterapia frente a enfermedades autoinmunes
-
Anticuerpos monoclonales en la terapia del cáncer
-
Otras estrategias anti-tumorales
-
Tratamiento “clásico” de la patología alérgica
-
Inmunoterapia en el trasplante de órganos
-
Tratamiento de inmunodeficiencias primarias
-
Vacunas (I): Bases inmunológicas
Expresión de las inmunoglobulinas en el BCR

Aquí tienes el temario explicado en el vídeo anterior. Si tienes alguna duda plantéala en el sistema de comentarios del final de la página.
Expresión de las inmunoglobulinas en el bcr
Exclusión alélica
Se denomina exclusión alélica al fenómeno por el cual tras la síntesis de una Ig funcional o productiva se detiene el reordenamiento (las recombinasas se paran y no se producen más bucles). Así cada célula B sólo tiene un tipo de Ac en su superficie o secretado. Esta especificidad se mantendrá durante toda la vida (únicamente podría sufrir cambios de afinidad, por mecanismos de hipermutación somática) del linfocito B. La exclusión alélica es, pues, responsable de la monoespecificidad para el reconocimiento del Ag por el linfocito B.
La célula B madura (que ya ha completado el reordenamiento genético de sus cadenas), en res- puestas primarias solo puede sintetizar IgM o IgD. Esto es por cuestiones físicas, (la longitud del mRNA alcanza a los genes Cμ y Cδ, que se encuentran los primeros en la secuencia del cromosoma 14). El que dicha inmunoglobulina sea de membrana o soluble dependerá de que en el procesamiento del RNA se incluyan o no los exones que codifican para la porción transmembranal y citoplasmática de la proteína o (alternativamente) de la porción secretora (Figura 8.11).
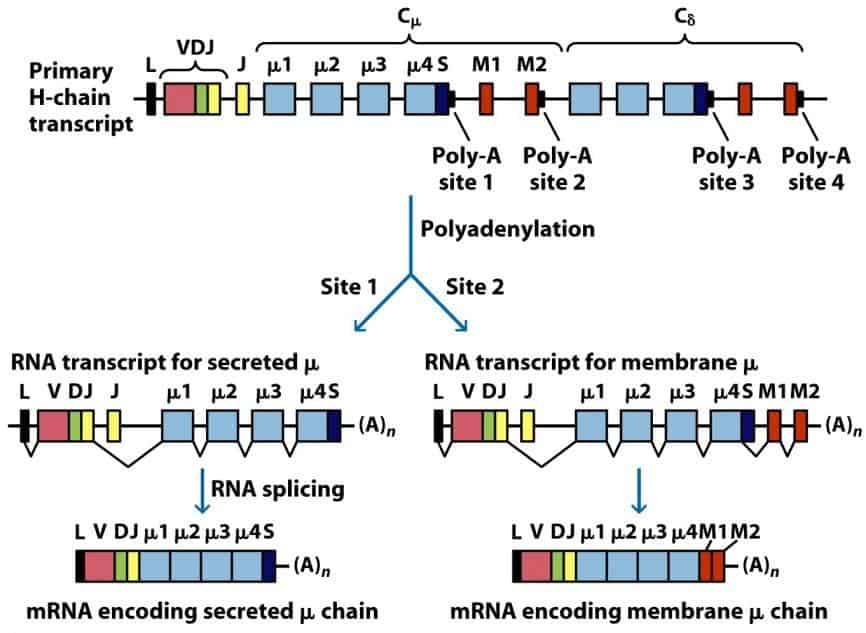
(Reproducido de Kindt T.J., Goldsby R.A., Osborne B.A. & Palacios R. (2007) Inmunología de Kuby (6ª Ed.) McGraw-Hill Interamericana, Madrid.)
La inmunoglobulina será IgM o IgD también dependiendo de dicho procesamiento del RNA (Figura 8.12). El resto de isotipos no se pueden producir inicialmente, ya que requieren cambios adicionales en el DNA. De modo que las inmunoglobulinas que puede producir un linfocito B maduro inicialmente son: mIgM, sIgM, mIgD, sIgD (donde s y m indican soluble o membrana).
Cambio de isotipo
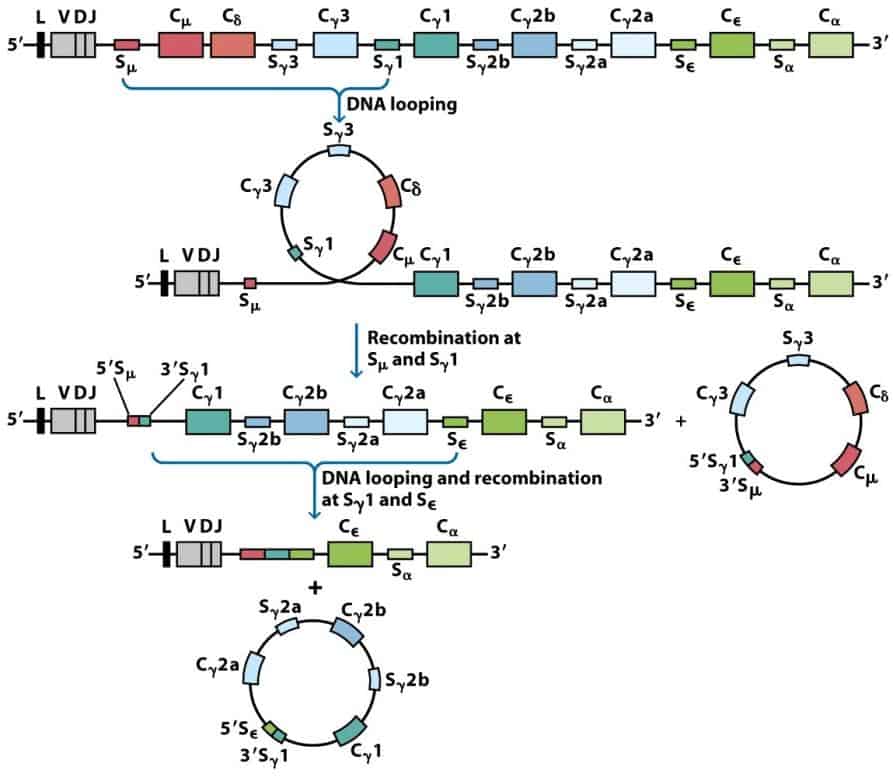
Para sintetizar otros isotipos tiene que producirse el denominado cambio de isotipo. El cambio de isotipo se produce varias veces en una línea clonal y no implica a las regiones variables (cuyo reordenamiento en médula ósea se mantiene siempre, por ejemplo V1D3J9), sino a los genes de las regiones constantes (Figura 8.13). Es muy frecuente en células B en la médula ósea.
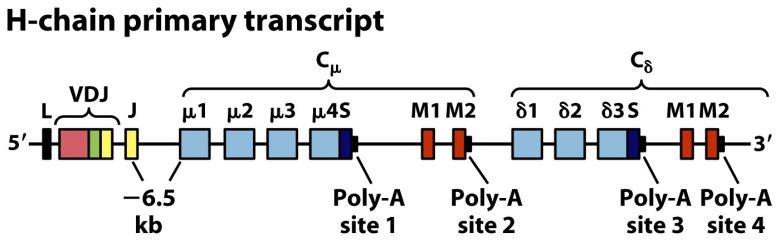
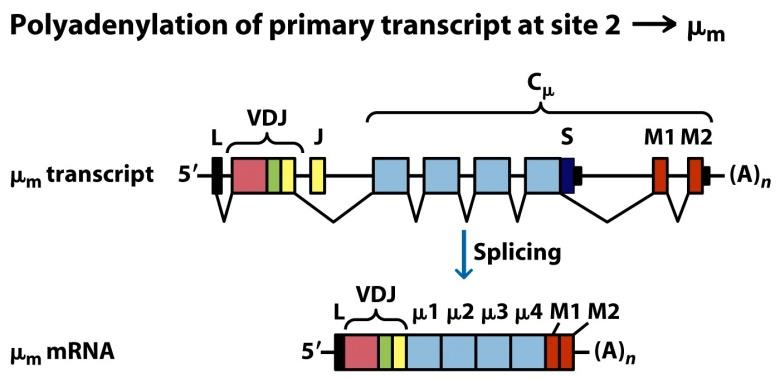
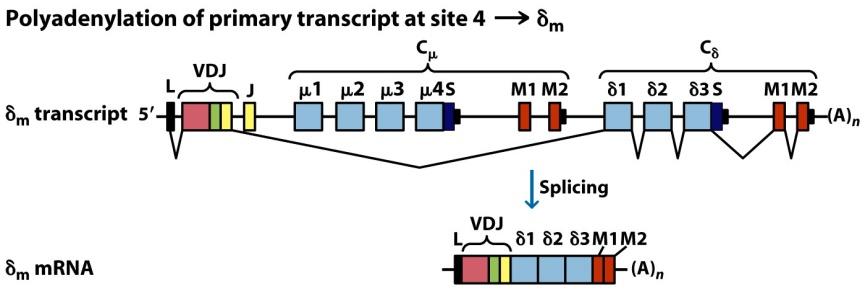
(Reproducido de Kindt T.J., Goldsby R.A., Osborne B.A. & Palacios R. (2007) Inmunología de Kuby (6ª Ed.) McGraw-Hill Interamericana, Madrid.)
(Reproducido de Kindt T.J., Goldsby R.A., Osborne B.A. & Palacios R. (2007) Inmunología de Kuby (6ª Ed.) McGraw-Hill Interamericana, Madrid.)
El cambio de isotipo, es un nuevo proceso de reordenamiento del ADN, y para que se produzca se necesitan dos requisitos indispensables:
- La participación de enzimas de recombinación.
- La presencia de secuencias nucleotídicas repetitivas denominadas “señales de cambio de clase”. Estas secuencias se encuentran delante de todos los genes que codifican para los fragmentos constantes, a excepción de IgD, y serán las que “guíen” el cambio, indicando donde tiene que producirse la recombinación.en el que se van perdiendo genes constantes de las diferentes cadenas pesadas.
El cambio de isotipo se puede producir múltiples veces en la vida de un clon, pero es irreversible (si se ha cambiado de IgM a IgE, esa célula jamás producirá IgM de nuevo). El límite lo pone cuando se cambia al isotipo IgA2 (puesto que el gen Cα2 es el último gen constante en el cromosoma 14).
El cambio de isotipo es inducido durante la cooperación entre Linfocitos B y T; son las células T quienes señalizan a las células B para que reordenen su DNA hacia nuevos segmentos constantes adyacentes a los segmentos VDJ. Estas células y su progenie cambian de secretar Ig M a otros isotipos. Para esto es preciso un contacto célula a célula entre T y B que esta mediado por el CD40L y CD40, respectivamente. Pero además, el linfocito Th ha de recibir dos señales de activación sucesivas: la 1ª se da cuando Th reconoce al complejo Ag+HLA de clase II (gracias a su receptor y a su correceptor, CD4), la 2ª es la unión de CD28 a su ligando, que es CD80 y CD86 en la célula B.
Después de que Th haya recibido estas dos señales, envía otras dos señales para el cambio de isotipo del linfocito B:
- Primera señal: expresión en superficie del ligando de CD40, que se unirá a CD40 en la superficie del linfocito B.
- Segunda señal: liberación de citocinas que se unirán a sus receptores en el linfocito B.
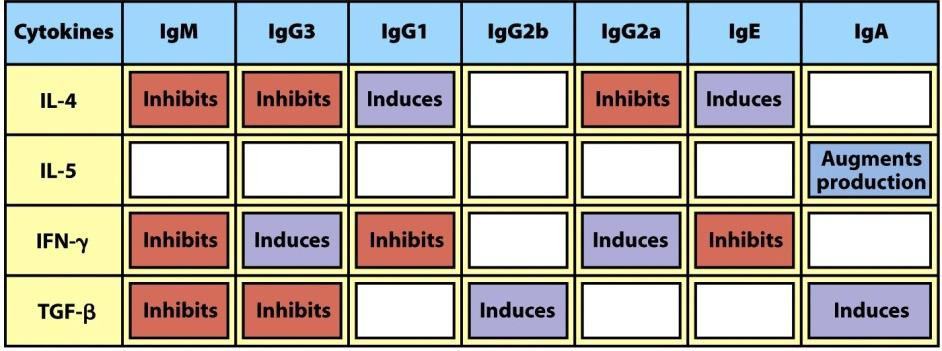
En función de la citocina liberada, se induce el cambio a diferentes isotipos (Figura 8.14):
- Interleucina-4: induce cambio a IgE.
- Interferón-γ: induce cambio a IgG.
- TGF-β (factor de crecimiento y transformación β): induce cambio a IgA.
- IL-4+IL-13: NO induce cambio de clase.
(Reproducido de Murphy K., Travers P. & Walport M. (2008) Janeway’s Immunobiology (7th Ed.) Garland Science, Nueva York.)
Responses