Inmunología Humana
-
INTRODUCCIÓN AL SISTEMA INMUNE HUMANOIntroducción. Conceptos básicos10 Temas
-
Introducción Inmunología
-
Funciones sistema inmune y barreras de defensa
-
Inmunidad innata vs adaptativa
-
Respuesta humoral vs celular
-
Respuesta adaptativa primaria vs secundaria
-
Características de la respuesta inmunitaria adaptativa
-
Concepto de antígeno
-
La respuesta inmunitaria específica es clonal
-
Revisión de los componentes del sistema inmunitario
-
Patología general del sistema inmunitario
-
Introducción Inmunología
-
Células del sistema inmune y diferenciación celular6 Temas
-
Hematopoyesis
-
Las células sanguíneas (I): Granulocitos y Mastocitos
-
Las células sanguíneas (II): Plaquetas, Eritrocitos, Monocitos y Macrófagos
-
Las células sanguíneas (III): Linfocitos (T y B), Células plasmáticas y LGL (Linfocitos NK)
-
Las células sanguíneas (IV): Células dendríticas y proporciones relativas de los tipos celulares
-
Funciones y gestión de receptores para antígenos
-
Hematopoyesis
-
Tejidos del sistema inmune: órganos linfoides 1º y 2º3 Temas
-
Células y mecanismos de la inmunidad innata (I): macrófagos, receptores y mecanismos efectores5 Temas
-
Células y mecanismos de la inmunidad innata (II): linfocitos NK, receptores y mecanismos efectores4 Temas
-
MOLÉCULAS IMPLICADAS EN EL RECONOCIMIENTO DE ANTÍGENOEl receptor de antígeno del linfocito B6 Temas
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
Inmunoglobulinas (II): Clases, formas, variabilidad y tejidos
-
Inmunoglobulinas (III): Enlaces antígeno-anticuerpo y funciones
-
Inmunoglobulinas (IV): Receptores para inmunoglobulinas
-
Inmunoglobulinas (V): Procesamiento de RNA, receptor del linfocito B, complejo co-receptor y activación célula B
-
Inmunoglobulinas (VI): Desarrollo / Diferenciación células B
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
El receptor de antígeno del linfocito T4 Temas
-
Mecanismos de generación de la diversidad de linfocitos T y B9 Temas
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
Reordenamiento de los genes
-
Mecanismos de amplificación de la diversidad
-
Expresión de las inmunoglobulinas en el BCR
-
Maduración de los linfocitos B
-
La cooperación T-B
-
Anomalías/Problemas en el proceso (CLÍNICA)
-
Maduración de los linfocitos B
-
Variabilidad en el repertorio del TcR en linfocitos T
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
El complejo principal de histocompatibilidad (I): estructura proteica, genética y nomenclatura3 Temas
-
El complejo principal de histocompatibilidad (II): Procesamiento y presentación de antígeno, polimorfismo y aplicaciones clínicas5 Temas
-
MOLÉCULAS ACCESORIAS DE LA RESPUESTA INMUNEEl sistema del complemento y sus receptores (I): vía clásica y vía alternativa4 Temas
-
El sistema del complemento y sus receptores (II): vía de las lectinas, vía lítica y regulación3 Temas
-
Moléculas implicadas en la comunicación intercelular (I): citocinas y sus receptores5 Temas
-
Moléculas implicadas en la comunicación intercelular (II): moléculas de adhesión y sus ligandos3 Temas
-
EL SISTEMA INMUNE EN ACCIÓN BLOQUEGeneración de linfocitos T efectores4 Temas
-
Generación de linfocitos B efectores7 Temas
-
Sistema Inmune asociado a mucosas (MALT)9 Temas
-
Introducción MALT
-
Estructura del tejido linfoide asociado a la mucosa intestinal
-
Recirculación y migración selectiva de linfocitos al intestino
-
Barrera epitelial intestinal: Función Inmunológica
-
Receptores para el reconocimiento de patrones moleculares
-
Inmunoglobulinas Poliméricas: IgM + IgA
-
Vías de entrada del antígeno en el intestino
-
Diferenciación de los linfocitos T efectores MALT
-
Tolerancia Oral ante antígenos solubles
-
Introducción MALT
-
La respuesta inmune (I): inmunidad innata e inflamación aguda8 Temas
-
Barreras no específicas frente a infección por microorganismos patógenos
-
Principales leucocitos que paticipan en las inmunidad innata
-
La rección inflamatoria
-
Inflamación aguda
-
Sucesos del proceso inflamatorio
-
Citocinas producidas por macrófagos: efectos locales y sistémicos
-
Efectos de las citocinas en la inmunidad innata
-
Proteínas de fase aguda
-
Barreras no específicas frente a infección por microorganismos patógenos
-
La respuesta inmune (II): mecanismos de la inmunidad específica8 Temas
-
La respuesta inmune (III): respuesta frente a virus, bacterias y hongos, protozoos y helmintos9 Temas
-
Principales enfermedades infecciosas mortales
-
Patógenos infecciosos
-
Respuestas frente a bacterias extracelulares
-
Respuesta frente a bacterias intracelulares
-
Proceso de fagocitosis
-
Respuestas frente a protozoos y helmintos
-
Ciclo vital del plasmodium
-
Respuestas inmunitarias frente a virus
-
Mecanismos de evasión inmunitaria. Virales y bacterianos.
-
Principales enfermedades infecciosas mortales
-
REGULACIÓN e INTRODUCCIÓN A LA INMUNOPATOLOGÍARegulación de la respuesta inmune (I): regulación por moléculas8 Temas
-
Regulación de la respuesta inmune: Introducción
-
Regulación por moléculas: el complemento (I)
-
Proteínas reguladoras del complemento y proceso de regulación (II)
-
Proceso de regulación por complemento (III)
-
Regulación de los anticuerpos preformados
-
Regulación por anticuerpos: Redes idiotípicas
-
Regulación por moléculas de linfocitos T
-
Regulación a nivel bioquímico: ITAM frente a ITIM
-
Regulación de la respuesta inmune: Introducción
-
Regulación de la respuesta inmune (II): regulación por células y sistemas4 Temas
-
El sistema inmune a lo largo del ciclo vital: Inmunosenescencia6 Temas
-
Introducción a la inmunopatología13 Temas
-
Conceptos básicos
-
Inmunodeficiencias
-
Las Inmunodeficiencias (II): Déficits de Complemento
-
Las Inmunodeficiencias (III): Déficits de citocinas en receptores
-
Las Inmunodeficiencias (IV): Déficits de TLR's
-
Las Inmunodeficiencias (V): Déficits de función fagocitaria
-
Las Inmunodeficiencias (VI): Déficit de linfocitos B
-
Las Inmunodeficiencias (VII): Déficits de linfocitos T
-
Inmunodeficiencias primarias (IDP's): Clínica y Diagnóstico
-
Hipersensibilidades
-
Autoinmunidad
-
Trasplantes. Rechazo
-
Inmunidad frente a tumores
-
Conceptos básicos
-
Introducción a la Inmunoterapia8 Temas
-
Vacunas (I): Bases inmunológicas
-
Vacunas (II): Tipos de vacunas
-
Inmunoterapia frente a enfermedades autoinmunes
-
Anticuerpos monoclonales en la terapia del cáncer
-
Otras estrategias anti-tumorales
-
Tratamiento “clásico” de la patología alérgica
-
Inmunoterapia en el trasplante de órganos
-
Tratamiento de inmunodeficiencias primarias
-
Vacunas (I): Bases inmunológicas
Presentación de antígeno (II): Ruta endocítica y tercera ruta

Aquí tienes el temario explicado en el vídeo anterior. Si tienes alguna duda plantéala en el sistema de comentarios del final de la página.
Vía de las moléculas MHCde clase II
Las moléculas MHC de clase II son heterodímeros formados por una cadena α y una cadena β unidas de forma no covalente. Ambas cadenas son sintetizadas de forma independiente en el retículo endoplasmático. Junto a ellas se sintetiza una proteína de cadena invariante (I), también llamada CD74, que estabiliza el heterodímero y tapona la cavidad de unión de péptidos.
En esta ruta de presentación, el patógeno es captado e internalizado a un endosoma (fagosoma) mediante fagocitosis por una célula presentadora de antígenos profesional. El fagosoma donde se ha integrado el patógeno se fusiona con un lisosoma primario cargado de enzimas hidrolíticas y proteasas (captesinas). La fusión de ambos da lugar a un lisosoma secundario o fagolisosoma en el que, tras la degradación del patógeno, aparecen péptidos antigénicos de distintas longitudes y secuencias. Los fagolisosomas vierten su contenido a un endosoma exclusivo de las células presentadoras de antígenos profesionales, denominado compartimento de carga de péptidos (Figura 10.3, flechas rojas).
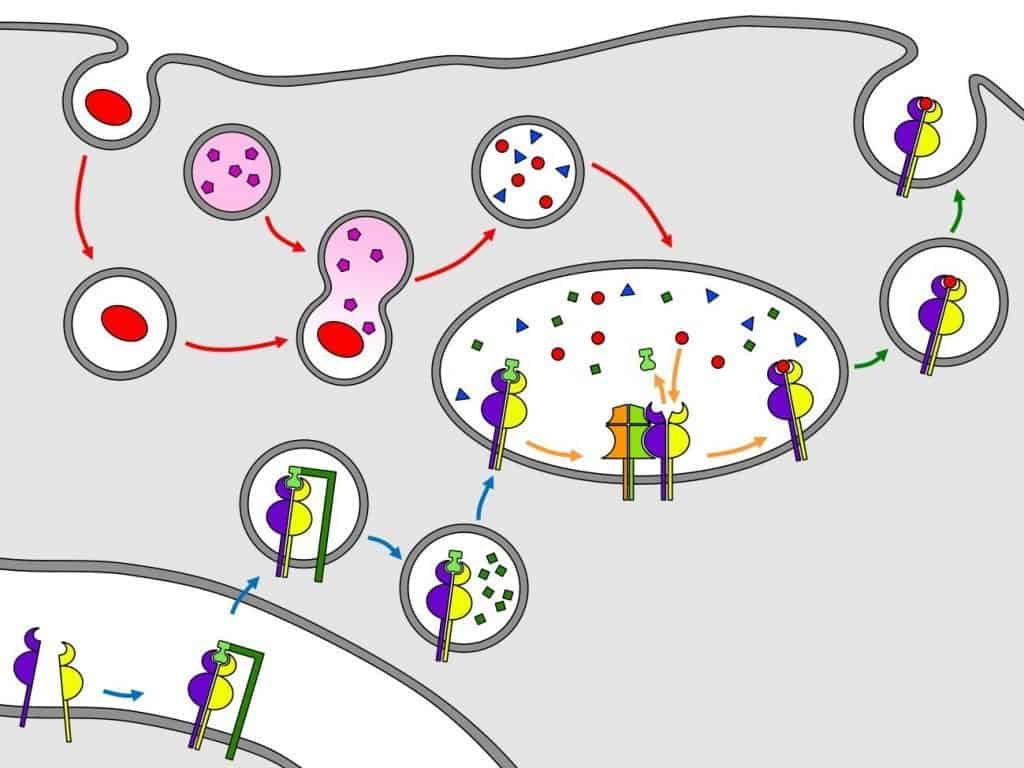
Los antígenos extracelulares son degradados por un fagolisosoma y enviados al compartimento de carga de péptidos. Las moléculas MHC de clase II sintetizadas en el retículo endoplasmático son muy inestables y, por ello, se asocian a CD74. MHC de clase II unido a CD74 se dirige al compar- timento de carga de péptidos y, en el transcurso, CD74 es hidrolizado salvo en una porción deno- minada CLIP, que tapona la cavidad de unión de péptidos. En el compartimento de carga de pépti- dos, HLA-DM permite la salida del CLIP y la entrada de uno de los péptidos antigénicos en MHC de clase II. La molécula MHC de clase II unida a su péptido es enviada al aparato de Golgi, donde sufre procesos de glicosilación. Por último, la molécula MHC de clase II unida a su péptido es enviada a la superfice de la célula presentadora de antígenos profesional. (Cortesía de David Pérez
Torres, Universidad de Valladolid.)
Las moléculas MHC de clase II taponadas por CD74 migran desde el retículo endoplasmático al compartimento de carga de péptidos, un orgánulo exclusivo de las células presentadoras de antígenos profesionales. Durante la migración, CD74 es hidrolizada salvo en la porción que tapona la cavidad de unión de péptidos, denominada CLIP (Figura 10.3, flechas azules).
En el compartimento de carga de péptidos se encuentran péptidos antigénicos procedentes de los fagolisosomas y moléculas MHC de clase II taponadas por el CLIP. Para que se puedan formar complejos antígeno-molécula MHC de clase II se requiere la participación de HLA-DM, una molécula que actúa como un intercambiador peptídico: expulsa el CLIP y expone la cavidad de unión de péptidos de la molécula MHC de clase II, permitiendo la unión de los péptidos antigénicos. Las moléculas HLA-DM difieren de las moléculas del MHC de clase II en que no son polimórficas, no se asocian con la cadena invariante (I) (CD74), y no se expresan en la superficie celular (Figura 10.3, flechas naranjas).
Cuando uno de los péptidos antigénicos se une a la cavidad de unión de péptidos para la que es específico, MHC de clase II completa su plegamiento y migra al aparato de Golgi, donde sufre procesos de glicosilación. Después de realizar las modificaciones necesarias, es enviada a la membrana de la célula presentadora de antígenos profesional en una vesícula de exocitosis (Figura 10.3, flechas verdes). El péptido alojado en la cavidad de unión de péptidos de la proteína MHC es presentado un receptor de la célula T que reconoce el péptido y las cadenas de MHC simultáneamente. Si no se reconocen ambas estructuras a la vez, no se produce la activación del linfocito T.
Responses