Inmunología Humana
-
INTRODUCCIÓN AL SISTEMA INMUNE HUMANOIntroducción. Conceptos básicos10 Temas
-
Introducción Inmunología
-
Funciones sistema inmune y barreras de defensa
-
Inmunidad innata vs adaptativa
-
Respuesta humoral vs celular
-
Respuesta adaptativa primaria vs secundaria
-
Características de la respuesta inmunitaria adaptativa
-
Concepto de antígeno
-
La respuesta inmunitaria específica es clonal
-
Revisión de los componentes del sistema inmunitario
-
Patología general del sistema inmunitario
-
Introducción Inmunología
-
Células del sistema inmune y diferenciación celular6 Temas
-
Hematopoyesis
-
Las células sanguíneas (I): Granulocitos y Mastocitos
-
Las células sanguíneas (II): Plaquetas, Eritrocitos, Monocitos y Macrófagos
-
Las células sanguíneas (III): Linfocitos (T y B), Células plasmáticas y LGL (Linfocitos NK)
-
Las células sanguíneas (IV): Células dendríticas y proporciones relativas de los tipos celulares
-
Funciones y gestión de receptores para antígenos
-
Hematopoyesis
-
Tejidos del sistema inmune: órganos linfoides 1º y 2º3 Temas
-
Células y mecanismos de la inmunidad innata (I): macrófagos, receptores y mecanismos efectores5 Temas
-
Células y mecanismos de la inmunidad innata (II): linfocitos NK, receptores y mecanismos efectores4 Temas
-
MOLÉCULAS IMPLICADAS EN EL RECONOCIMIENTO DE ANTÍGENOEl receptor de antígeno del linfocito B6 Temas
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
Inmunoglobulinas (II): Clases, formas, variabilidad y tejidos
-
Inmunoglobulinas (III): Enlaces antígeno-anticuerpo y funciones
-
Inmunoglobulinas (IV): Receptores para inmunoglobulinas
-
Inmunoglobulinas (V): Procesamiento de RNA, receptor del linfocito B, complejo co-receptor y activación célula B
-
Inmunoglobulinas (VI): Desarrollo / Diferenciación células B
-
Inmunoglobulinas (I): Formas en la naturaleza, estructura y dominio básico
-
El receptor de antígeno del linfocito T4 Temas
-
Mecanismos de generación de la diversidad de linfocitos T y B9 Temas
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
Reordenamiento de los genes
-
Mecanismos de amplificación de la diversidad
-
Expresión de las inmunoglobulinas en el BCR
-
Maduración de los linfocitos B
-
La cooperación T-B
-
Anomalías/Problemas en el proceso (CLÍNICA)
-
Maduración de los linfocitos B
-
Variabilidad en el repertorio del TcR en linfocitos T
-
Genes para las cadenas pesadas y ligeras: Diversidad potencial
-
El complejo principal de histocompatibilidad (I): estructura proteica, genética y nomenclatura3 Temas
-
El complejo principal de histocompatibilidad (II): Procesamiento y presentación de antígeno, polimorfismo y aplicaciones clínicas5 Temas
-
MOLÉCULAS ACCESORIAS DE LA RESPUESTA INMUNEEl sistema del complemento y sus receptores (I): vía clásica y vía alternativa4 Temas
-
El sistema del complemento y sus receptores (II): vía de las lectinas, vía lítica y regulación3 Temas
-
Moléculas implicadas en la comunicación intercelular (I): citocinas y sus receptores5 Temas
-
Moléculas implicadas en la comunicación intercelular (II): moléculas de adhesión y sus ligandos3 Temas
-
EL SISTEMA INMUNE EN ACCIÓN BLOQUEGeneración de linfocitos T efectores4 Temas
-
Generación de linfocitos B efectores7 Temas
-
Sistema Inmune asociado a mucosas (MALT)9 Temas
-
Introducción MALT
-
Estructura del tejido linfoide asociado a la mucosa intestinal
-
Recirculación y migración selectiva de linfocitos al intestino
-
Barrera epitelial intestinal: Función Inmunológica
-
Receptores para el reconocimiento de patrones moleculares
-
Inmunoglobulinas Poliméricas: IgM + IgA
-
Vías de entrada del antígeno en el intestino
-
Diferenciación de los linfocitos T efectores MALT
-
Tolerancia Oral ante antígenos solubles
-
Introducción MALT
-
La respuesta inmune (I): inmunidad innata e inflamación aguda8 Temas
-
Barreras no específicas frente a infección por microorganismos patógenos
-
Principales leucocitos que paticipan en las inmunidad innata
-
La rección inflamatoria
-
Inflamación aguda
-
Sucesos del proceso inflamatorio
-
Citocinas producidas por macrófagos: efectos locales y sistémicos
-
Efectos de las citocinas en la inmunidad innata
-
Proteínas de fase aguda
-
Barreras no específicas frente a infección por microorganismos patógenos
-
La respuesta inmune (II): mecanismos de la inmunidad específica8 Temas
-
La respuesta inmune (III): respuesta frente a virus, bacterias y hongos, protozoos y helmintos9 Temas
-
Principales enfermedades infecciosas mortales
-
Patógenos infecciosos
-
Respuestas frente a bacterias extracelulares
-
Respuesta frente a bacterias intracelulares
-
Proceso de fagocitosis
-
Respuestas frente a protozoos y helmintos
-
Ciclo vital del plasmodium
-
Respuestas inmunitarias frente a virus
-
Mecanismos de evasión inmunitaria. Virales y bacterianos.
-
Principales enfermedades infecciosas mortales
-
REGULACIÓN e INTRODUCCIÓN A LA INMUNOPATOLOGÍARegulación de la respuesta inmune (I): regulación por moléculas8 Temas
-
Regulación de la respuesta inmune: Introducción
-
Regulación por moléculas: el complemento (I)
-
Proteínas reguladoras del complemento y proceso de regulación (II)
-
Proceso de regulación por complemento (III)
-
Regulación de los anticuerpos preformados
-
Regulación por anticuerpos: Redes idiotípicas
-
Regulación por moléculas de linfocitos T
-
Regulación a nivel bioquímico: ITAM frente a ITIM
-
Regulación de la respuesta inmune: Introducción
-
Regulación de la respuesta inmune (II): regulación por células y sistemas4 Temas
-
El sistema inmune a lo largo del ciclo vital: Inmunosenescencia6 Temas
-
Introducción a la inmunopatología13 Temas
-
Conceptos básicos
-
Inmunodeficiencias
-
Las Inmunodeficiencias (II): Déficits de Complemento
-
Las Inmunodeficiencias (III): Déficits de citocinas en receptores
-
Las Inmunodeficiencias (IV): Déficits de TLR's
-
Las Inmunodeficiencias (V): Déficits de función fagocitaria
-
Las Inmunodeficiencias (VI): Déficit de linfocitos B
-
Las Inmunodeficiencias (VII): Déficits de linfocitos T
-
Inmunodeficiencias primarias (IDP's): Clínica y Diagnóstico
-
Hipersensibilidades
-
Autoinmunidad
-
Trasplantes. Rechazo
-
Inmunidad frente a tumores
-
Conceptos básicos
-
Introducción a la Inmunoterapia8 Temas
-
Vacunas (I): Bases inmunológicas
-
Vacunas (II): Tipos de vacunas
-
Inmunoterapia frente a enfermedades autoinmunes
-
Anticuerpos monoclonales en la terapia del cáncer
-
Otras estrategias anti-tumorales
-
Tratamiento “clásico” de la patología alérgica
-
Inmunoterapia en el trasplante de órganos
-
Tratamiento de inmunodeficiencias primarias
-
Vacunas (I): Bases inmunológicas
Introducción y las 3 vías del complemento

Aquí tienes el temario explicado en el vídeo anterior. Si tienes alguna duda plantéala en el sistema de comentarios del final de la página.
Conceptos básicos y nomenclatura
Cuando un patógeno atraviesa la barrera epitelial y franquea la primera línea de defensa antimicrobiana del individuo, el primer componente al que éste se enfrenta es un elemento de la inmunidad innata conocido como sistema de complemento, o simplemente, complemento.
El complemento es una colección de proteínas solubles presentes en la sangre y los fluidos corporales. Existen más de 30 proteínas que pertenecen al complemento y su producción es funda- mentalmente hepática. En ausencia de infección, estas proteínas circulan en forma inactiva. En presencia de patógenos o de anticuerpos unidos a patógenos, las proteínas del sistema de complemento se activan.
Las proteínas del complemento interaccionan entre sí de maneras concretas a través de distintas vías de activación, cuya consecuencia final es la muerte del patógeno. Este objetivo se alcanza mediante la facilitación de la fagocitosis y la inducción de una respuesta inflamatoria capaz de combatir la infección. Existen tres vías de activación del complemento: la vía clásica, que se inicia gracias a los anticuerpos adheridos a la superficie del patógeno, la vía alternativa, que se inicia por la simple presencia del antígeno, y la vía de las lectinas, que se activa por moléculas de la superficie del patógeno (Figura 11.1). Todas ellas convergen en la formación de la conver- tasa de C3.
Las proteínas del sistema de complemento se sintetizan en el hígado en forma de zimógenos o proenzimas, es decir en forma inactiva, y requieren un procedimiento de escisión proteolítica para activarse. Con carácter general, cada proteína del sistema de complemento es activada por otra proteína del sistema. Las cascadas de proteólisis así generadas son el origen de las vías de activación, y son las responsables de la generación de los componentes efectores del complemento (vía final común) y de la ampliación de la respuesta, que permiten eliminar al patógeno.
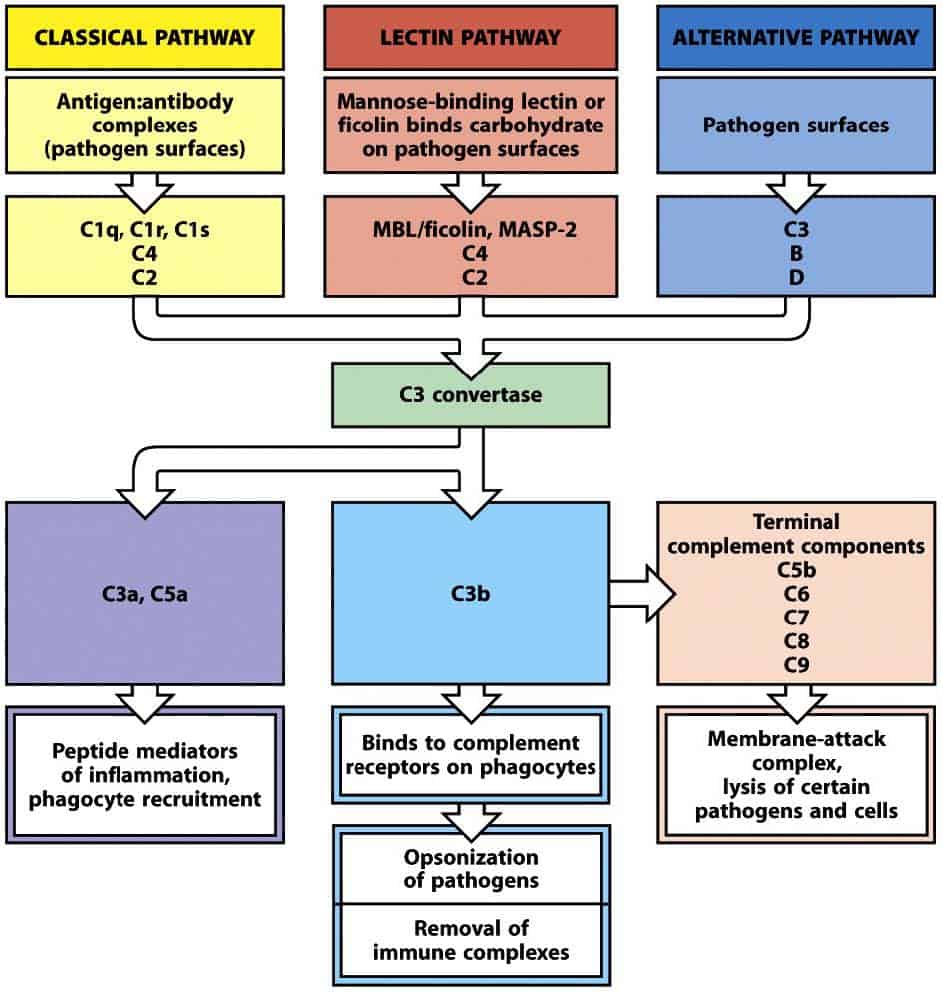
(Reproducido de Murphy K et al. (2012) Janeway’s Immunobiology (8th Ed.) Garland Science, Nueva York.)
Nomenclatura
La nomenclatura de las proteínas del sistema de complemento puede resultar confusa. La primera vía de activación del complemento descubierta fue la vía clásica, cuyas proteínas se de- signan con la letra C y un número (C1, C2, C3, C4, C5, C6, C7, C8 y C9). Dicho número no describe el orden en el que interaccionan los elementos, sino el orden en que las moléculas fue- ron descubiertas. Cuando una proteasa escinde una proteína del complemento para activarla, da lugar a dos productos. Los productos de esta reacción se designan con sufijo formado por una letra minúscula. El fragmento menor de la reacción proteolítica se denomina anafilotoxina, se designa con la letra a (C3a, C5a,…), permanece soluble en el medio y tiene función quimiotáctica para las células inflamatorias. El fragmento mayor de la reacción proteolítica se denomina convertasa, se designa con la letra b (C3b, C5b,…), se deposita sobre la superficie del patógeno y tiene actividad serinproteasa para actuar sobre la siguiente proteína del complemento inactiva. Existen dos excepciones a esta nomenclatura general. La primera es la correspondiente a la proteólisis de C2, cuyo fragmento mayor se denominó C2a inicialmente por ser la fracción activa- da, y cuya nomenclatura se ha mantenido durante mucho tiempo. No obstante, en la actualidad es preferible denominar C2b a este fragmento. La segunda es la nomenclatura de C1q, C1r y C1s, que no son productos de la proteólisis de C1, sino un grupo de proteínas que se asocian para formar C1.
Las proteínas de la vía alternativa fueron descubiertas más tarde y de designaron con el término factor y una letra mayúscula (factor B, factor D…). Los productos de su proteólisis se designan igual que los de la vía clásica, con las letras minúsculas a, para el fragmento menor (Ba,…), y b, para el fragmento mayor (Bb,…).
La vía clásica
La vía clásica del complemento es similar a la vía de las lectinas, pero se inicia por un sensor de patógenos denominado complejo C1 o, simplemente, C1. C1 puede interaccionar directamente con algunos patógenos, pero también puede interaccionar con anticuerpos. Por tanto, la vía clásica tiene una función en la inmunidad innata y también en la inmunidad adaptativa.
Al igual que el complejo MBL-MASP, el complejo C1 está formado por una subunidad grande, denominada C1q, que actúa como sensor de patógenos, y dos serinproteasas, denominadas C1r y C1s (Figura 11.4). C1q es un hexámero de trímeros, formados por monómeros que contienen un dominio globular aminoterminal y un dominio de tipo colágeno carboxiterminal. Los trímeros se ensamblan mediante interacciones entre los dominios de tipo colágeno, de manera que los dominios globulares se unen formando una cabeza globular. La molécula completa de C1q tiene seis cabezas globulares unidas por las colas tipo colágeno, que permiten reconocer los patóge- nos o anticuerpos unidos a su superficie, especialmente IgG e IgM. C1r y C1s se relacionan estrechamente con MASP-2, mientras que MASP-1 se relaciona con ellas de forma menos importante. C1r y C1s interaccionan de forma no covalente para formar pares C1r:C1s. Los pares se unen a los brazos de C1q, quedando una parte de estos complejos en posición externa a C1q.
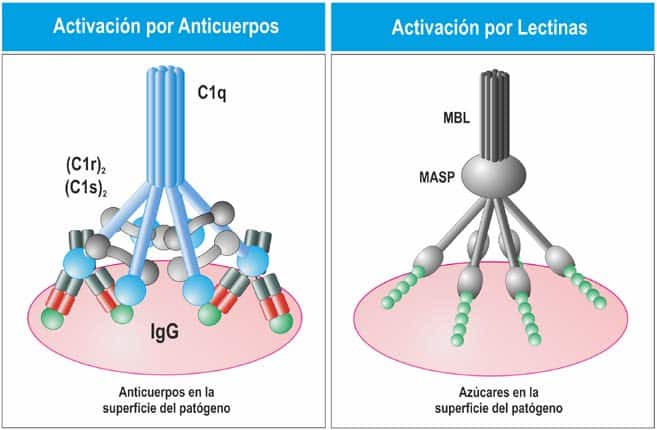
(Reproducido de Regueiro J.R., López C., González S. & Martínez E. (2011) Inmunología. Biolo- gía y Patología del Sistema Inmune. (4ª Ed.) Editorial Médica Panamericana, Madrid. Reproduci-
do de Murphy K et al. (2012) Janeway’s Immunobiology (8th Ed.) Garland Science, Nueva York.)
La unión de dos o más cabezas de C1q a su ligando produce un cambio de conformación en el complejo C1r:C1s, produciendo la activación de la actividad autocatalítica de C1r. Entonces, C1r hidroliza su proteína C1s asociada para generar una serinproteasas activa. C1s actúa en los siguientes dos componentes de la vía clásica, C4 y C2. C1s hidroliza C4 para producir C4b, que se une covalentemente a la superficie del patógeno, de la misma forma que en la vía de las lectinas. A C4b se une una molécula de C2, que es hidrolizada por C1s, liberándose la seriproteasa C2b. Este proceso da lugar a la formación de C4b2b, que es la convertasa de C3 de la vía clásica y de las lectinas (Figura 11.5 y Figura 11.6).
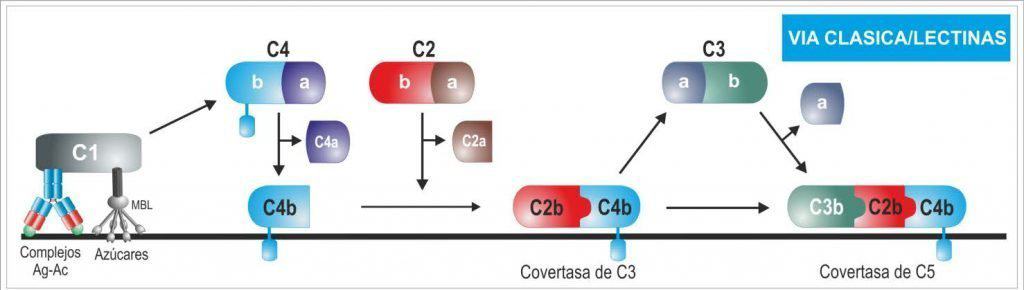
(Reproducido de Regueiro J.R., López C., González S. & Martínez E. (2011) Inmunología. Biología y Patología del Sistema Inmune. (4ª Ed.) Editorial Médica Panamericana, Madrid.)
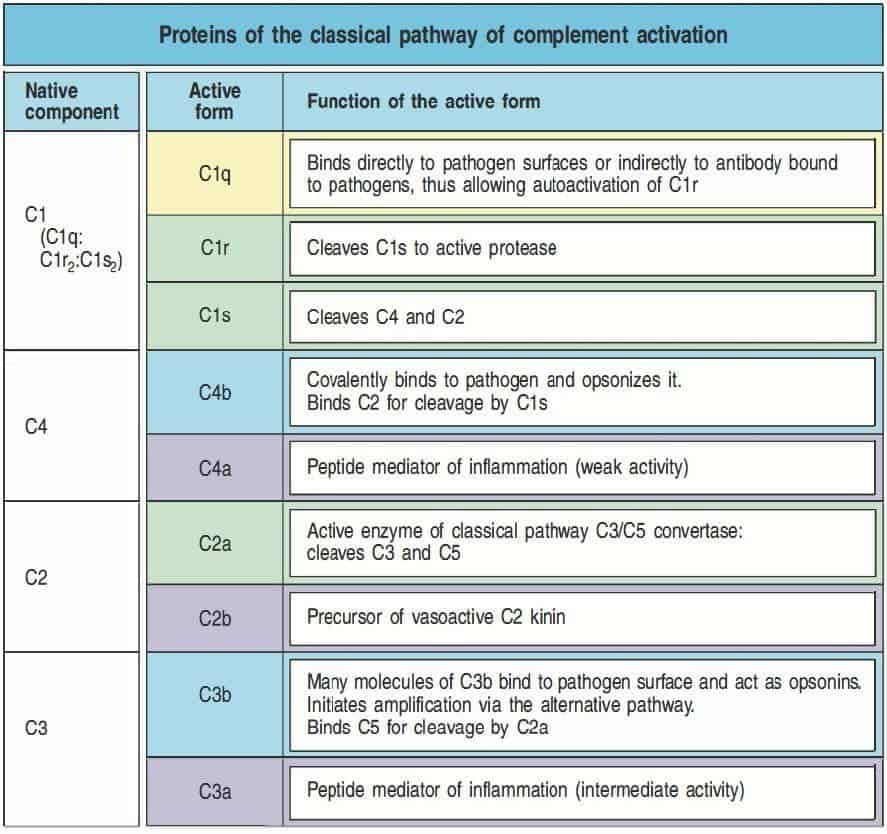
(Reproducido de Murphy K et al. (2012) Janeway’s Immunobiology (8th Ed.) Garland Science, Nueva York.)
Responses